Rapport de baguage 2005
Répartition des causes d'échec
|
|
|
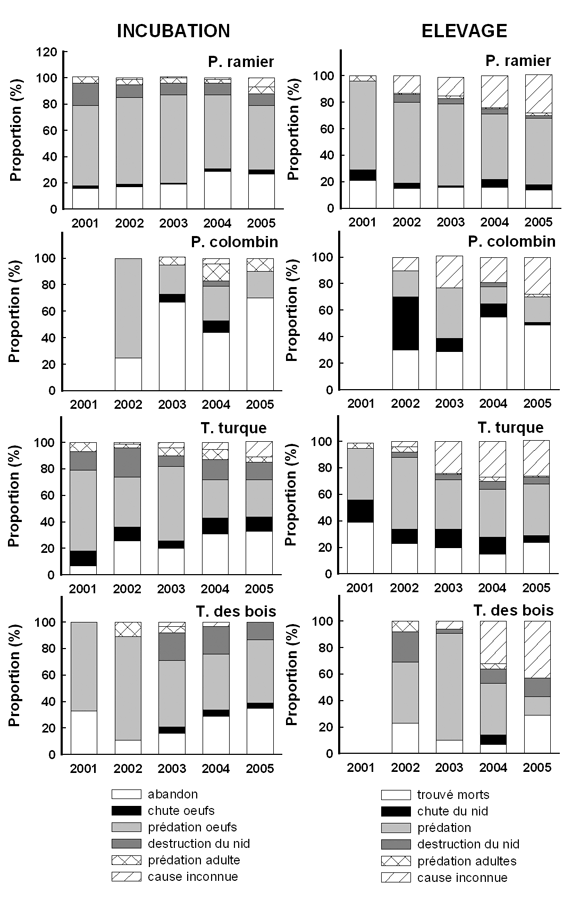
Fig. 11 : répartition des échecs selon les stades sur la période 2001 à 2005 ; incubation (noir), élevage (gris), stade inconnu (hachuré). |
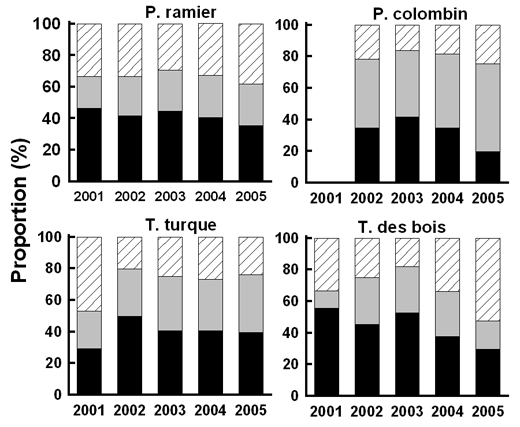
Si l’on considère que les échecs à un stade inconnu peuvent être répartis à parité entre l’incubation et l’élevage, on obtient une proportion des échecs pendants l’incubation (Fig. 11) toujours supérieure à 50% chez les 2 tourterelles et le P. ramier (maximum 59%). Les proportions sont inversées chez le P. colombin, avec des échecs survenant majoritairement pendant l’élevage (de 50 à 68% !). La mise en perspective des résultats obtenus depuis 2001 suggère un tassement des échecs en période d’incubation pour les 2 pigeons et la T. des bois. Ce tassement est d’au minimum de 10% chez le P. ramier et au maximum 25% chez la T. des bois, le P. colombin se retrouvant en position intermédiaire. Chez le P. Colombin, ce recul se fait au profit des échecs survenus pendant l’élevage, qui augmentent de 12%. En revanche chez les 2 autres espèces, ce recul des échecs sur œuf est en partie compensé par les échecs survenus à un stade inconnu, il est donc difficile de dire si ce recul est réel ou bien tout simplement lié au fait qu’il a été plus difficile d’identifier les cas d’échecs en incubation. Seule la T. turque montre une relative augmentation de la proportion d’échecs pendant l’incubation (20% entre 2001 et 2002 puis stagnation au delà).
On note également une certaine évolution dans les causes des échecs au cours de l’incubation et de l’élevage. Pendant l’incubation, la prédation des œufs reste la principale cause d’échec mais dans une proportion chaque année moins importante, au point même qu’elle se retrouve chez le P. colombin nettement derrière les échecs liés aux abandons de couvée. Les abandons de couvées sont d’ailleurs en proportion la 2ème cause d’échec pendant l’incubation et montrent une nette tendance à l’augmentation depuis le début du programme. Nous manquons cependant de précision sur la part de ces abandons qui concernent des couvées viables ou des œufs infertiles. Toutes les autres causes d’échec pendant l’incubation restent relativement minoritaires et dépassent très rarement 20% des échecs.
 Pendant l’élevage, le pattern est relativement similaire entre les espèces, à l’exception du P. colombin. La prédation des poussins reste la principale cause d’échec (elle a pu dépasser 80% chez la T. des bois !), quoiqu’en recul régulier depuis le début de ce programme. En revanche chez le P. colombin, la cause d’échec la plus couramment observée est la mort des poussins au nid, sans trace de prédation. Une 3ème catégorie se détache ensuite pour toutes les espèces, qui relève plus d’un constat que d’une quelconque explication : il s’agit d’échec pour raison inconnue, c’est à dire typiquement un nid vide de poussins, sans aucun indice pour fournir un diagnostic. On peut raisonnablement penser que la prédation assure une part importante de ces échecs mais sans que l’on puisse en apporter la preuve (les poussins ont également pu tomber du nid…).
Pendant l’élevage, le pattern est relativement similaire entre les espèces, à l’exception du P. colombin. La prédation des poussins reste la principale cause d’échec (elle a pu dépasser 80% chez la T. des bois !), quoiqu’en recul régulier depuis le début de ce programme. En revanche chez le P. colombin, la cause d’échec la plus couramment observée est la mort des poussins au nid, sans trace de prédation. Une 3ème catégorie se détache ensuite pour toutes les espèces, qui relève plus d’un constat que d’une quelconque explication : il s’agit d’échec pour raison inconnue, c’est à dire typiquement un nid vide de poussins, sans aucun indice pour fournir un diagnostic. On peut raisonnablement penser que la prédation assure une part importante de ces échecs mais sans que l’on puisse en apporter la preuve (les poussins ont également pu tomber du nid…).
Il faut remarquer que le recul de la prédation peut illustrer la réalité mais également résulter d’une plus grande rigueur dans les diagnostics réalisés par les observateurs, qui au début du programme pouvaient encore classer par erreur un nid vide d’œufs ou de poussins comme échec par prédation sans pourtant avoir d’indice.
|
Fig. 12 & 13 : volution sur la période 2001-2005 de la répartition (en %) des principales causes d’échec pendant l’incubation (à gauche) et l’élevage (à droite) |
|
|